
Vous avez sans doute déjà piqué un sprint pour attraper votre bus/votre métro. Dans ce genre de situation, vous avez conscience du fait que vous devez parcourir une certaine distance le plus vite possible pour atteindre votre objectif : vous arrêter pile devant les portes et vous engouffrer dans le véhicule. Des chercheurs du MIT montrent dans une nouvelle étude que cette expérience, en apparence anodine, est en réalité guidée par toute une série de calculs dans notre cerveau.
S’arrêter brusquement devant une cible après une course rapide n’est pas qu’un simple réflexe : cette capacité à passer brusquement d’un comportement à un autre vient du cortex cérébral, qui utilise les règles de vie apprises lors d’expériences antérieures pour élaborer et guider nos actions, expliquent les chercheurs. « Le but est l’endroit où le cortex intervient. Où suis-je censé m’arrêter pour atteindre mon objectif (monter dans le bus) », souligne dans un communiqué Mriganka Sur, professeur de neurosciences au Picower Institute for Learning and Memory du MIT et co-auteur de l’étude. Pour étudier en détail le contrôle de ce type de mouvements, Sur et son équipe ont réalisé des expériences sur des souris.
L’un de ses collègues et premier auteur de l’étude, Elie Adam, a élaboré des modèles mathématiques du comportement qui ont montré qu’un signal « d’arrêt » allant directement de la région M2 du cortex (le cortex moteur secondaire) à des régions du tronc cérébral des souris (qui contrôlent leurs pattes) serait traité trop lentement. « M2 envoie un signal d’arrêt, mais lorsque vous le modélisez et que vous faites appel aux mathématiques, vous constatez que ce signal, par lui-même, ne serait pas assez rapide pour que l’animal s’arrête à temps », explique Adam, premier auteur d’un nouvel article portant sur cette recherche, publié dans la revue Cell Reports. Une autre voie de signalisation, plus rapide, était donc forcément à l’œuvre.
Un arrêt brusque induit par deux signaux distincts
Dans le cadre de cette étude, les chercheurs ont développé une tâche dans laquelle des souris devaient courir sur un tapis roulant, s’arrêter au niveau d’un point de repère visuel (une série de LED défilant sur les côtés du tapis), puis s’arrêter et maintenir leur position pendant 1,5 seconde pour récupérer une récompense (un peu d’eau). En cas d’échec (dépassement du point de repère), un son était émis. Ils ont alors examiné la façon dont le cortex moteur secondaire (M2) contrôlait la locomotion à mesure des entraînements.
Lorsque l’on marche ou que l’on court sans but précis, le cerveau envoie « simplement » des signaux aux neurones de la moelle épinière, qui relayent ensuite ces signaux aux motoneurones qui contrôlent les muscles des jambes. Chaque signal marche/arrêt déclenche un pic d’activité électrique. Dès lors qu’un objectif est introduit (courir pour attraper un ballon ou son bus, ou une récompense ici), la voie de signalisation est différente.

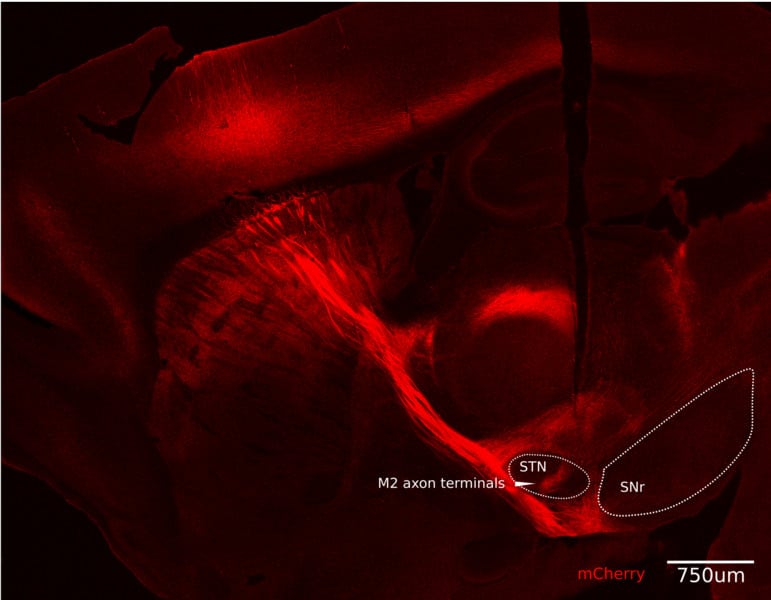 La coloration rouge met en évidence les projections axonales du cortex moteur M2. Celles qui mènent au noyau sous-thalamique (NST) sont particulièrement importantes. © E. Adam et al.
La coloration rouge met en évidence les projections axonales du cortex moteur M2. Celles qui mènent au noyau sous-thalamique (NST) sont particulièrement importantes. © E. Adam et al.Les chercheurs ont découvert que le M2 envoie bien un signal d’arrêt, mais à une région intermédiaire appelée noyau sous-thalamique (NST), qui elle-même envoie ensuite deux signaux — l’un inhibiteur, l’autre excitateur — par deux voies distinctes qui se rejoignent dans le tronc cérébral. La différence entre ces deux signaux, qui arrivent presque immédiatement, mais l’un après l’autre, entraîne une reconnaissance directe du changement — et donc une mise en œuvre beaucoup plus rapide de l’arrêt du mouvement. « Une poussée inhibitrice suivie d’une excitation peut créer un [changement de] signal aigu », explique Sur.
Plus la différence est grande, plus le changement initié par le signal inhibiteur est rapide et plus l’arrêt est brusque. Ce système de signalisation, qui s’apparente à un calcul de dérivée, est plus complexe, mais permet de contrôler les mouvements de façon plus précise. « Cela permet d’anticiper et de prévoir. Si je connais la dérivée, le taux de changement de vitesse, alors je peux prédire quelle sera ma vitesse à l’étape suivante », a déclaré à Quanta Magazine Sridevi Sarma, neuroscientifique à l’Université Johns Hopkins.
Une inhibition accentuée par l’excitation
L’expérience a confirmé que le M2 produisait une augmentation de l’activité neuronale, et envoyait des signaux vers le NST, uniquement lorsque les souris avaient besoin d’atteindre l’objectif pour lequel elles avaient été entraînées. « Ces signaux sont guidés visuellement et ne se produisent pas lors des arrêts de locomotion spontanés », note l’équipe dans Cell Reports. En outre, l’activation artificielle de la voie M2-NST obligeait les souris à s’arrêter, tandis que son inhibition artificielle faisait que les souris dépassaient le point de repère un peu plus souvent.
Les signaux inhibiteur et excitateur relayés par le NST au tronc cérébral parviennent plus exactement au noyau pédonculopontin (NPP), situé dans la région locomotrice mésencéphalique. Les scientifiques ont découvert que différents types de cellules du NPP répondaient avec un timing différent aux signaux. Avant l’arrêt, les cellules excitatrices étaient actives et reflétaient la vitesse de l’animal ; au moment de l’arrêt, ils ont observé deux types de poussées d’activité, l’une légèrement plus lente que l’autre, qui étaient transmises soit directement au NPP par la voie excitatrice, soit indirectement via une région appelée « substantia nigra pars reticulata » par la voie inhibitrice.


« Le résultat net de l’interaction de ces signaux dans le NPP était une inhibition accentuée par l’excitation », résume l’équipe. Cette étude corrobore d’autres travaux récents, dans lesquels Elie Adam a développé un modèle de la façon dont la stimulation cérébrale profonde à haute fréquence dans le NST pouvait corriger rapidement les problèmes moteurs résultant de la maladie de Parkinson.
Une autre étude a montré comment le cerveau supplante les voies de communication utilisées lors de mouvements réflexes dès qu’un contrôle exécutif est nécessaire. « Ces études permettent de comprendre comment le cortex peut contrôler consciemment des comportements moteurs instinctifs, mais aussi l’importance de régions plus profondes, telles que le NST, pour mettre rapidement en œuvre un comportement dirigé vers un but », résume l’équipe du Picower Institute for Learning and Memory.
Les chercheurs émettent l’hypothèse que le rôle de cette voie hyperdirecte pourrait s’étendre au-delà des processus moteurs, aux processus associatifs et cognitifs (telles que les interruptions et les changements de pensée ou d’humeur). Des recherches sont en cours pour éclaircir ce point.